C单倍群在整个东亚地区有着广泛的分布,在中国男性人口中的占比在10%以上,对于这么一个重要的单倍群其来源目前却依然略显神秘,而网络上也流传着很多说法,其中也有不少误解。本文试图对一些常见的说法做出解释。
问题一:C单倍群是老亚洲/早亚洲人吗?
答:首先,我们要了解“新老亚洲”概念提出的历史背景。在分子人类学这门学科发展之初,经历了很长时间的摸索的过程,在这个过程中出现了很多的猜测,有的被后来的研究逐步证实,而有的却逐渐显得站不住脚。
在研究者最早绘制的树形中,C、D、E、F是四个并列的支系,这使得很多人望文生义的认为C和D的关系更近,或者C和A、B那样是一个代表着众多“古老”支系的旁系群。但随着研究的深入,树形的完善。这四个单倍群的关系也逐渐清晰。实际上,这四个单倍群来自于同一个共同的祖先CT-M168,而这个祖先繁衍出了两个支系CF和DE。所以,相对于D和E单倍群,C和O的祖先F有着更近的关系。从这点上来看说C抛下更亲的兄弟F,而与关系更远的兄弟支系D先走出非洲,或者先来到东亚,就显得不太站得住脚了。
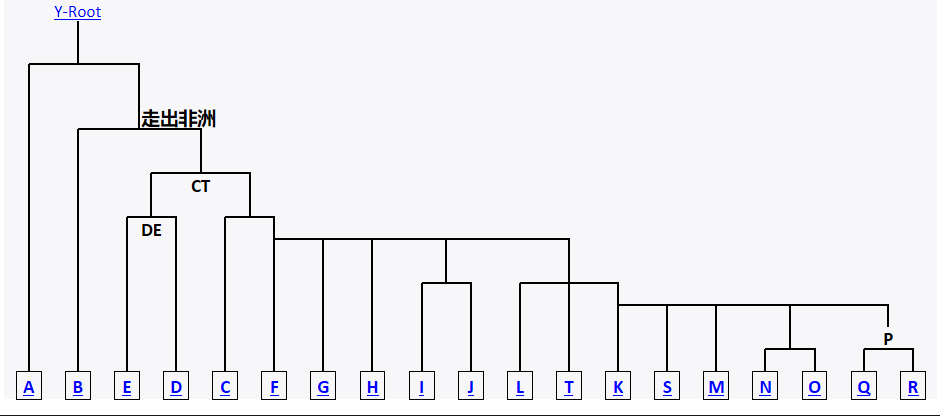
图一、 修正后的Y染色体支系树
而C单倍群作为“老亚洲”的另一个曾经的证据在于有考古证据表明澳大利亚在五六万年前就已经有了智人活动的踪迹。而C单倍群作为现今澳洲土著中占比最高的类型,自然成为了这批现代人父系最为可能的候选,如果C单倍群五六万年前就到达了澳洲,必然是历经东南亚而来的。相对于当时认为共祖时间只有两万多年的O来说,顺理成章的成为了“老亚洲”。然而随着现代研究结果的不断丰富,这一推理链条中的众多重要环节却出现了严重的断裂。第一,对于澳洲蒙戈湖古人DNA的研究显示,其母系的线粒体DNA不属于已知的现代人世系。可能是属于早于七万年前更早走出非洲的一批智人的后代,那么与F共祖时间不到七万年的C自然不可能在那时就已出现。第二,通过大样本量高通量数据的累积,我们发现整个C单倍群(包括澳洲样本)的共祖时间可能还不到五万年,只有四万八千年左右。澳洲的C单倍群C1b2b同当地的K*一样,同其他地区的兄弟支系分开都不超过45000年。如果说澳洲的C单倍群支系是五六万年前就已抵达的,那么广布于整个欧亚大陆的C单倍群就成了全部从澳大利亚分化而来的,这显然与我们在考古上的认识有着严重的矛盾。第三,NO单倍群在三万七千年前左右分化,这个分化地点极有可能在东亚及其周边地区,同样,C2单倍群在三万五千年前左右分化,分化地点也应是在东亚及周边地区。所以C2在东亚出现的时间未必会比NO更早,两者很有可能是在同一时期进入东亚地区,并共同参与了东亚人群的形成。
问题二:C单倍群是“棕种人”吗?
至于这个说法的来源,最初是因为在澳大利亚土著中C1单倍群占有较高的比例,澳洲土著既然一般被人为是“棕种人”,那么有人便把整个C单倍群都定义为棕种人。然而实际上,澳大利亚土著父系中C单倍群的比例只有一半左右,还有近一半的比例的父系属于NORQ的上游K单倍群的一个支系。而澳洲无论C单倍群还是K单倍群都是与其他在大陆的近亲分化了四万年以上的支系。也就是说澳洲土著父系中的C1和K很可能是在四万年前结伴进入澳洲的。如果仅仅因为澳洲土著父系中有C而将整个C单倍群定义为“棕种人”,那么在同样的逻辑下K单倍群包括其下游的NORQ也同样会被定义为“棕种人”。
另一方面来说,目前我们对于“人种”的定义是来基于相貌、肤色这些体质上的特征,这些体质上的特征也正是基因外在的体现。Y染色体在整个基因组中所在的比例不到1%,在Y染色体上目前尚未发现与体质外貌相关的基因。所以与“人种”所关联的遗传信息基本上都来源于常染色体,而非Y染色体。如果说Y染色体能决定人种或种族,那么没有Y染色体的女性又要怎么定位呢,所以这种思路本身就带有一种极度男权的色彩。
此外,用肤色来定义人种,是因为肤色往往是我们视觉上最为直观的外貌特征,然而,目前所知的几种浅肤色突变SCL24A5(西欧亚常见)、OCAII(东亚常见)出现的时间可能都非常晚,研究者一般认为在两万年之内。在这些浅肤色突变出现之前,我们的祖先可能都是一种较深肤色的状态。即使抛却肤色这点来说,西班牙发现的中石器时代猎人La Brana古DNA检测显示父系为C1-V20,但从常染上分析,最接近现代的北欧人群,而且还有着蓝色的眼睛。所以仅凭Y染难以构成判断种族的标准的。

问题三:C3、C2傻傻分不清楚。
这主要是由于Y染色体支系树系统命名变化导致的。在研究的初期,研究者发现C单倍群下有几个明显的分支,便命名为C1(常见于日本)、C2(常见于太平洋群岛)、C3(常见于东亚、北亚、美洲)、C4(常见于澳大利亚)、C5(常见于南亚、西亚)、C6(常见于欧洲),然而随着研究的深入,研究者们发现,C1、C2、C4、C5、C6存在共享的突变,也就是有着一个更近的共同祖先,C3则是与他们更早分开的一支。所以在新的系统命名中将原C3外的几支合并为新的C1,原C3便被命名为新的C2。

图三、C单倍群系统命名新老对照表(复旦严实)
问题四:C2(原C3)是蒙古人/通古斯人/阿尔泰人吗?
因为C2(原C3)高频出现于北亚地区的民族中,比如蒙古族、哈萨克族、赫哲族、鄂伦春族等,而这些民族与阿尔泰/满-通古斯语系有着非常密切的联系,所以不少人认为C2单倍群是与阿尔泰/通古斯语系民族相关的。但事实并非这么简单。
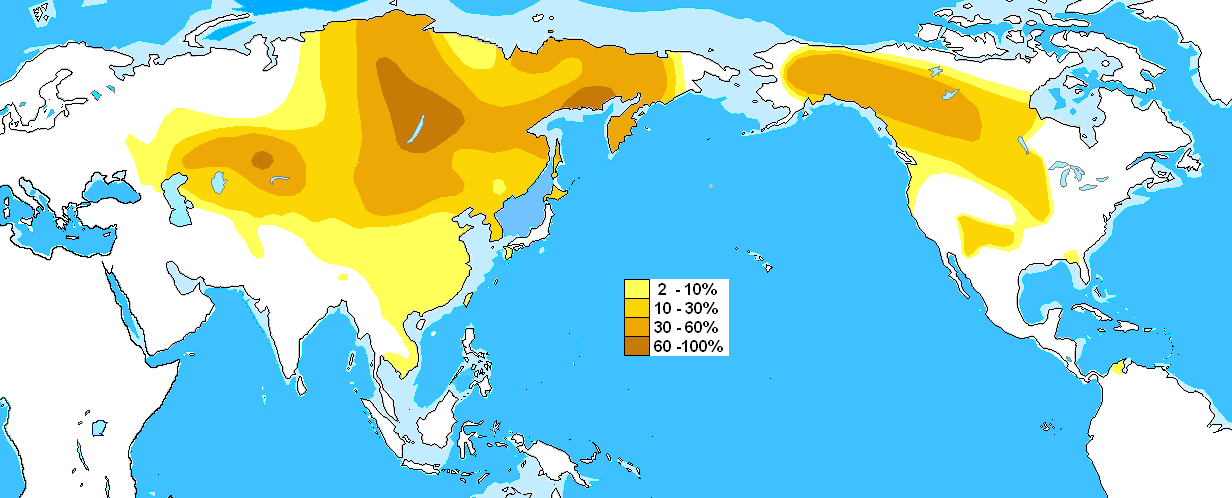
图四、单倍群C2-M217分布图
整个C2单倍群的共祖时间高达三万五千年,也就是说在三万五千年前,C2单倍群分化为L1396和F1067两个大的支系,即我们平时所说的C2北支和C2南支。这个分化的时间要远远早于我们目前所知的语系或民族所形成的时间。C2北支确实在蒙古、鄂伦春、赫哲、哈萨克等民族中有着较高频率的分布。不仅如此,其下游的一支P39还在一万年前左右进入了美洲,在北美印第安人,特别是纳德内语族的部落中有着高频的分布。
而C2南支在分布上则与C2北支在地域上有着很大的差异,C2南支主要分布在汉族当中,占到整个汉族人口的10%左右,是汉族人群的五个超级祖先之一。此外在朝鲜、日本、土家族、侗族、苗族、白族等汉族周边的地区和民族里也有着广泛的分布。相比之下C2北支在汉族的分布区只有不到1%左右。
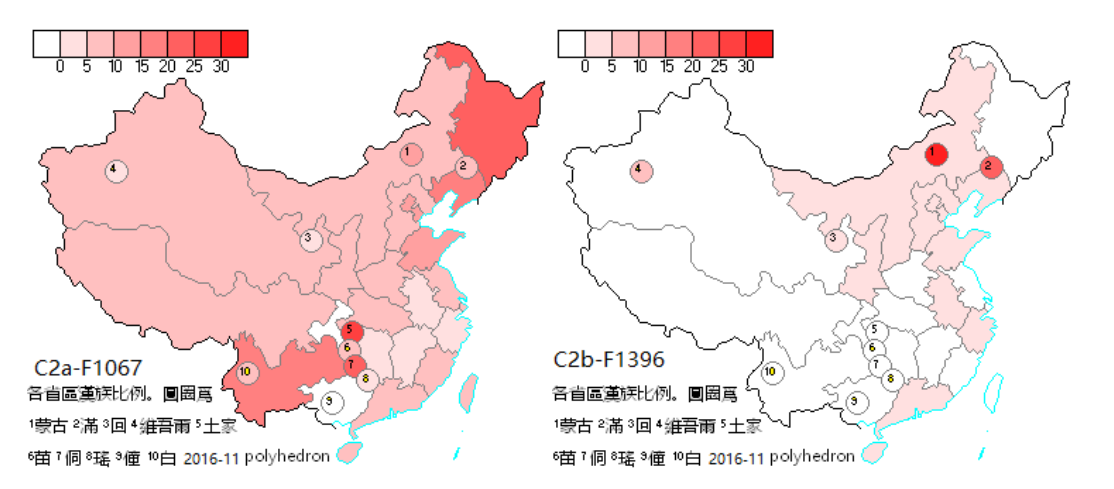
图五、Y染色体单倍群C2a(南支)与C2b(北支)全国分布图(复旦严实)
省份区域为汉族,圆圈为标注的少数民族
总而言之,相对于C2、O、N这些共祖时间在两三万年以上的单倍群来说,现代民族的形成时间是非常短暂,可能只有几千甚至几百年。在这些民族形成之前,不同父系单倍群的人群可能早就已经交融生活在了一起。也正如不断出现的考古结果所展示给我们的那样,正是史前时代以中原为中心的多个文化中心的交流碰撞才共同孕育了波澜壮阔的中华文明。
平南陈
点赞
莊
9 赞